Abstract
Background. Diabetes mellitus (DM) is a complex condition, with salivary gland malfunction being one of its complications. The management of the disease presents numerous side effects. Consequently, botanical products have been proposed as a promising alternative for the development of safe and effective treatments.
Objectives. The aim of the study was to investigate the antioxidant activity of chia seeds (Salvia hispanica L.) as a potential therapeutic tool for alleviating the deleterious effects of DM in albino rats.
Material and methods. The study sample comprised 66 male albino rats. Group I (n = 6) received a saline solution, group II (n = 30) was subjected to diabetic induction via a single intraperitoneal injection of streptozotocin (STZ), and group III (n = 30) was subjected to diabetic induction and treated with 250 mg/kg/day of powdered chia seeds for 2 weeks prior to the administration of STZ and until the end of the study. The rats were euthanized 45 days after the induction of DM. The parotid and mandibular mucosal glands were extracted and prepared for histological (hematoxylin and eosin (H&E)), immunohistochemical (vascular endothelial growth factor (VEGF)) and comet assay analyses.
Results. Marked degenerative changes were evident in the acini and duct architecture of the parotid and mandibular mucosal glands in group II. These findings were confirmed histologically, immunohistochemically and via the comet assay. In contrast, in group III, treatment with chia seeds resulted in the regeneration of salivary gland tissues.
Conclusions. Chia seeds demonstrated antioxidant properties against DM-induced salivary gland dysfunction in albino rats.
Keywords: diabetes mellitus, regeneration, Salvia hispanica L., salivary glands, antioxidant properties
Introduction
Diabetes mellitus (DM) is a metabolic condition characterized by high blood glucose levels and disruption of carbohydrate, lipid and protein metabolism.1 It is a disorder of the endocrine system in which a decrease in the production of insulin results in elevated blood glucose levels.2 In addition, uncontrolled DM results in end organ damage affecting the genitourinary, cardiovascular and neurological systems.3
Salivary glands are a set of major and minor glands that secrete saliva into the oral cavity through the duct system. Rats have multiple minor glands and 3 pairs of major salivary glands, namely the parotid, submandibular and sublingual glands.4
The production of saliva is necessary to maintain the equilibrium of the oral cavity and overall health.5 In diabetic conditions, salivary glands are exposed to deleterious effects, leading to reduced saliva production, destruction of the periodontium, salivary hypofunction, and oral mucous membrane injury.6 An association has been established between complications of diabetes and salivary malfunction. Xerostomia (dry mouth due to a deficient amount of saliva) and polydipsia (pathological thirst) are frequently observed in diabetic subjects as a result of diminished salivary secretion.7 Many authors have stated that the decreased salivary flow rate in diabetic patients is caused by increased urination, which leads to a reduction in extracellular fluid volume, necessary for saliva production by the salivary glands.8
Recent studies have demonstrated that hyperglycemia occurs as a consequence of DM. The condition produces oxidative and cellular impairments, which in turn negatively affect the normal structure of organs and tissues.9 Constant elevation of blood sugar levels leads to long-term oxidative stress that triggers the excessive production of reactive oxygen species (ROS). The occurrence of diabetic complications and their progression are mediated by ROS.10 Reactive oxygen species are highly reactive chemical molecules that, when significantly elevated under stressful conditions, lead to profound damage to cellular structures, especially when antioxidant systems are compromised.11
Various oxidative stress markers, such as 8-hydroxy-2’-deoxyguanosine (8-OHdG), protein carbonyl (PC), 4-hydroxynonenal–protein adduct (4-HNE), oxidized and/or malondialdehyde (MDA)-modified low-density lipoprotein (LDL) cholesterol (oxy-LDL/MDA-LDL) and 8-isoprostanes (8-isoP), have been investigated in streptozotocin (STZ)-induced DM rats to assess oxidative damage in the salivary glands. The results indicated that the 3 major salivary glands are subjected to increased oxidative stress regardless of disease duration. However, the parotid glands showed the greatest extent and diversity of oxidative injury.11
At present, salivary gland dysfunction is treated by stimulation of salivary secretion, either through oral receptors using ascorbic acid, by stimulating mastication (e.g., chewing gum), or by pharmacological stimulation using pilocarpine. Nevertheless, all of these remedies cause several complications and provide only short-time relief; therefore, effective long-term therapy is still needed.12
Another treatment approach for DM relies on oral hypoglycemic agents or insulin. Besides being costly, these drugs cause several side effects, such as decreased blood sodium levels, jaundice, nausea, headaches, vomiting, and increased body weight.13 In low-, middle- and high-income countries, populations often rely on botanical products for health maintenance, because they are frequently considered a safe and effective approach to managing chronic diseases. Lately, a variety of botanical products have been accepted for the treatment of DM and its complications.14
Over the last few years, medicinal plants have emerged as promising therapeutic agents with hypoglycemic effects for DM treatment.15 Shukla et al. examined a variety of herbal remedies for the treatment of DM, including aloe vera, eucalyptus, garlic, and ginger.16 Chia (Salvia hispanica L.), which belongs to the genus Salvia,17 is a plant native to northern Guatemala and southern Mexico. It was used as a nutritional food source by pre-Columbian populations.18 Chia is distinguished by its high content of alpha-linolenic acid (omega-3), fiber, proteins, vitamins, minerals, and phytonutrients, including phenolic compounds.19 These constituents contribute to improvements in biological markers associated with non-communicable diseases, exhibiting anti-inflammatory,20 antioxidant,21 hypotensive,22 hypocholesterolemic,23 and hypoglycemic properties.24
Chia may assist in the prevention of cardiovascular diseases, inflammatory and neurological disorders, as well as diabetes, as stated by Vuksan et al., who proposed that a high-fiber diet may contribute to the management of diabetes.25 The study was carried out on 20 patients with diabetes, who were given bread made with chia flour and additional whole seeds to be sprinkled on foods at home. The total consumption of chia seeds was 37 g/day. The study results demonstrated a small reduction in blood glucose as well as reduced levels of systemic inflammation, measured by C-reactive protein (CRP). Notably, blood became less susceptible to coagulation, which constitutes a risk factor that may lead to heart attacks and strokes.25
Objectives
This study aimed to assess the therapeutic efficacy of chia seeds in ameliorating salivary gland dysfunction caused by DM in diabetic rats.
Material and methods
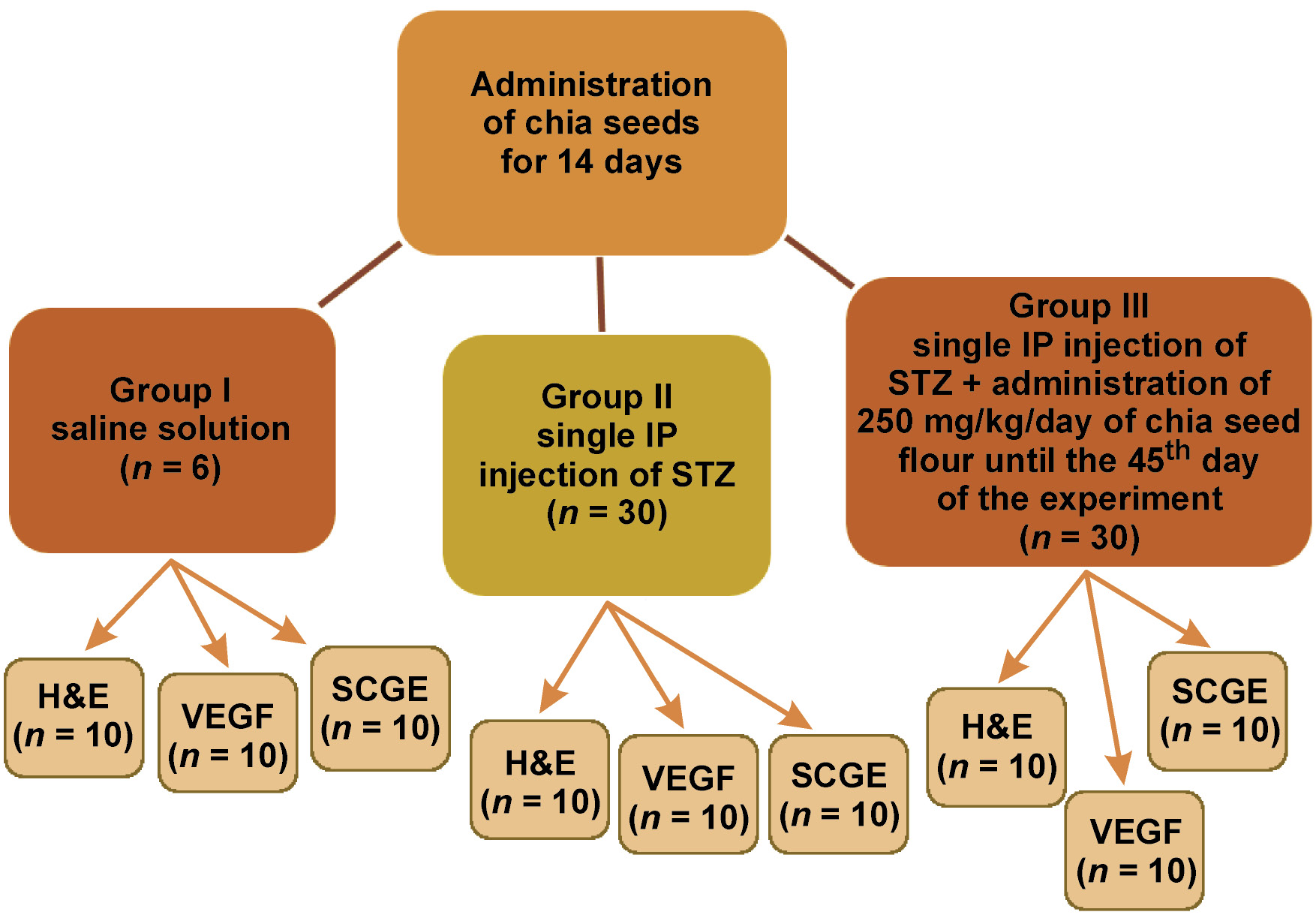
A total of 66 mature male albino rats, weighing 200–250 g, were utilized in the current research. The animals were housed in separate cages and provided with a standard rodent diet and water. Prior to the study, the rats were administered chia seeds for 14 days. The animals were divided into the following 3 groups:
• group I (control group; n = 6) – administered a saline solution;
• group II (n = 30) – subjected to diabetic induction via a single intraperitoneal injection of STZ without additional treatment;
• group III (n = 30) – subjected to diabetic induction via a single intraperitoneal injection of STZ and treated with 250 mg/kg/day of chia seed flour for 2 weeks prior to diabetic induction and until the end of the study.26
The study design is presented in Figure 1.
Parotid and mandibular mucosal gland assessment
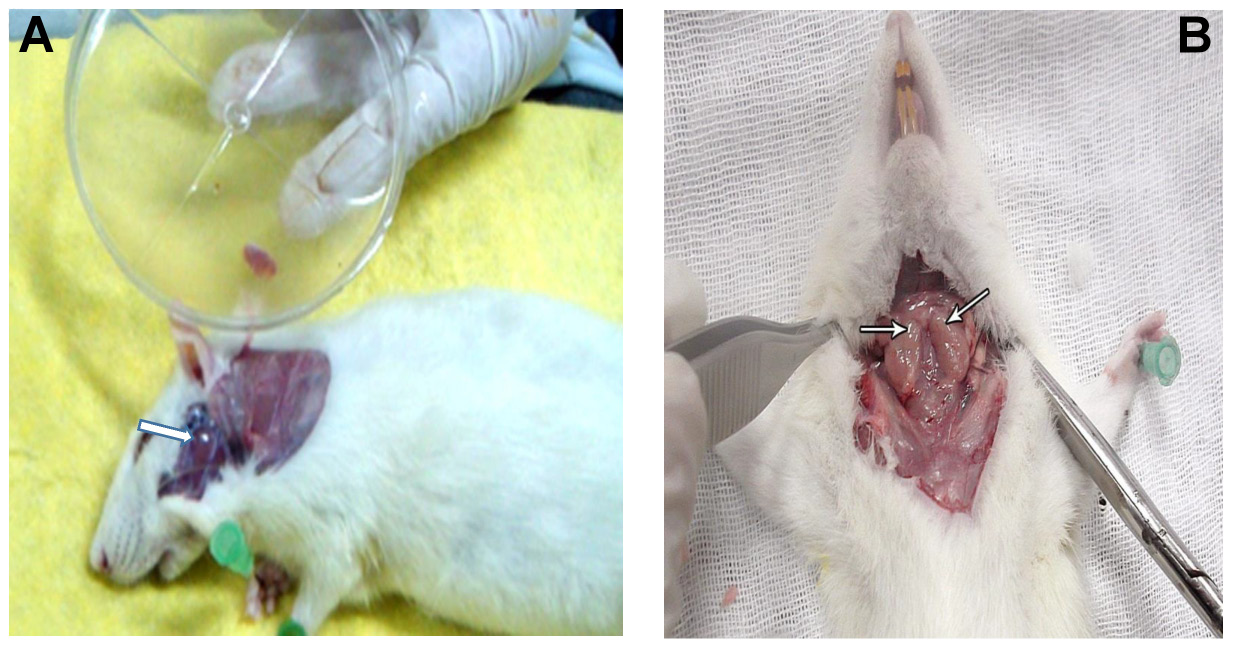
The rats were euthanized by an overdose of halothane 45 days after diabetic induction.27 The parotid and mandibular mucosal glands were removed (Figure 2) and divided as follows:
• 2 rats from group I, 10 rats from group II and 10 rats from group III were used for histological examination by hematoxylin and eosin (H&E) staining. The excised glands were fixed in 10% neutral-buffered formalin for 24 h, immersed in paraffin and sectioned at 5 μm;
• 2 rats from group I, 10 rats from group II and 10 rats from group III were used for immunohistochemical examination of vascular endothelial growth factor (VEGF). The excised glands were preserved in 10% neutral-buffered formalin for 24 h, immersed in paraffin and sectioned at 5 μm. Subsequently, they were immunostained using the avidin–biotin method. The sections were stained with rabbit anti-rat VEGF polyclonal antibodies (Sigma-Aldrich, St. Louis, USA);
• 2 rats from group I, 10 rats from group II and 10 rats from group III were used for single-cell gel electrophoresis (comet assay). The excised glands were stored in 1 mL of saline solution in Eppendorf tubes at −180°C.28, 29
Histological staining
Hematoxylin and eosin staining was performed as a routine histological procedure. For microscopic analysis, the excised glands were fixed in 10% formalin for 24 h. The specimens were washed thoroughly under running water, dehydrated through a graded series of alcohol, cleared in xylene, and embedded in paraffin wax according to standard techniques.30 After embedding in paraffin, 5 μm-thick sections were cut and stained with H&E.
These histological sections were investigated under a light microscope (Olympus CH2; Olympus Optical Co., Ltd., Tokyo, Japan).
Immunohistochemical evaluation
The sectioned glands were subjected to immunostaining using the avidin–biotin technique. The sections were stained with rabbit anti-rat VEGF polyclonal antibodies (Sigma-Aldrich).
Five non-overlapping fields from each slide were digitized using a digital camera (Olympus Optical Co., Ltd.) mounted on a microscope (Olympus CH2). The images were analyzed on an Intel Core i3-based computer using VideoTesT-Morphology software (Video Test, Saint Petersburg, Russia) with a specific built-in routine for immunostain quantification. The targeted, positively immunostained areas were automatically selected and separated from the original image based on the stain hue range. The selected area was thresholded and defined as a region of interest. Subsequently, a 3D histogram was constructed, and the integrated density of the area was calculated.31
Diabetes induction
Experimental diabetes was induced in rats from groups II and III after overnight fasting32 by a single intraperitoneal injection of STZ (STZ: 45 mg/kg body weight in 0.1 mL citrate buffer, pH 4.5). Three days after the injection, the presence of diabetes was confirmed by hyperglycemia (non-fasting blood glucose level >300 mg/dL) using a blood glucose meter.33 The blood glucose levels were monitored at 3 and 45 days after induction, and the results are presented in Table 1.
Comet assay
The comet assay,28, 29 also referred to as single-cell gel electrophoresis, is a rapid and highly sensitive fluorescence microscopy-based technique for the assessment of DNA damage and repair in individual nucleated cells. The method is based on the measurement of the ability of denatured, cleaved DNA fragments to migrate out of the cell under the influence of an electric field, whereas undamaged DNA migrates more slowly and remains within the nucleoid. The evaluation of the DNA “comet” tail shape and migration pattern allows for the assessment of DNA damage. In this assay, cells are immobilized in a layer of low-melting-point agarose, followed by gentle cell lysis, and treated with alkaline solution to unwind and denature the DNA and hydrolyze sites of damage. The samples are then submitted to electrophoresis, stained with a fluorescent DNA-intercalating dye, and visualized using epifluorescence microscopy.35
Chia administration
Group III received a daily dose of 250 mg/kg of chia seeds (PLANTON, New Delhi, India).26 The doses used in animal studies vary from 3.0% to 41.3% for chia seeds, and from 0.15% to 10.00% for chia oil, as reported by Enes et al.34 Freshly prepared seeds were minced and immersed in water 1 day prior to administration via a stomach tube. The administration was commenced 2 weeks before the induction of diabetes and continued until the end of the experiment.26
Statistical analysis
The data was analyzed using the IBM SPSS Statistics for Windows software, v. 20.0 (IBM Corp., Armonk, USA). The results are presented as mean ± standard deviation (M ±SD). Differences between the groups were assessed using one-way analysis of variance (ANOVA). Subsequently, a post hoc least significant difference (LSD) test was performed. Values were considered statistically significant at p < 0.05.
The DNA fragmentation pattern was estimated using a computer-assisted image analysis system (Comet Assay IV™; Perceptive Instruments). A total of 100 haphazardly selected cells per slide were recorded. DNA damage was assessed by evaluating the percentage of DNA in the comet tail relative to the total cellular DNA content.
Results
Clinical observations
A total of 8 rats were excluded from the study, as 5 rats died within 24 h after diabetes induction, and in group II, 2 rats died 1 week after diabetes induction, and 1 rat died after the 2nd week of diabetes induction. The rats were replaced to maintain group sizes.
Histological evaluation
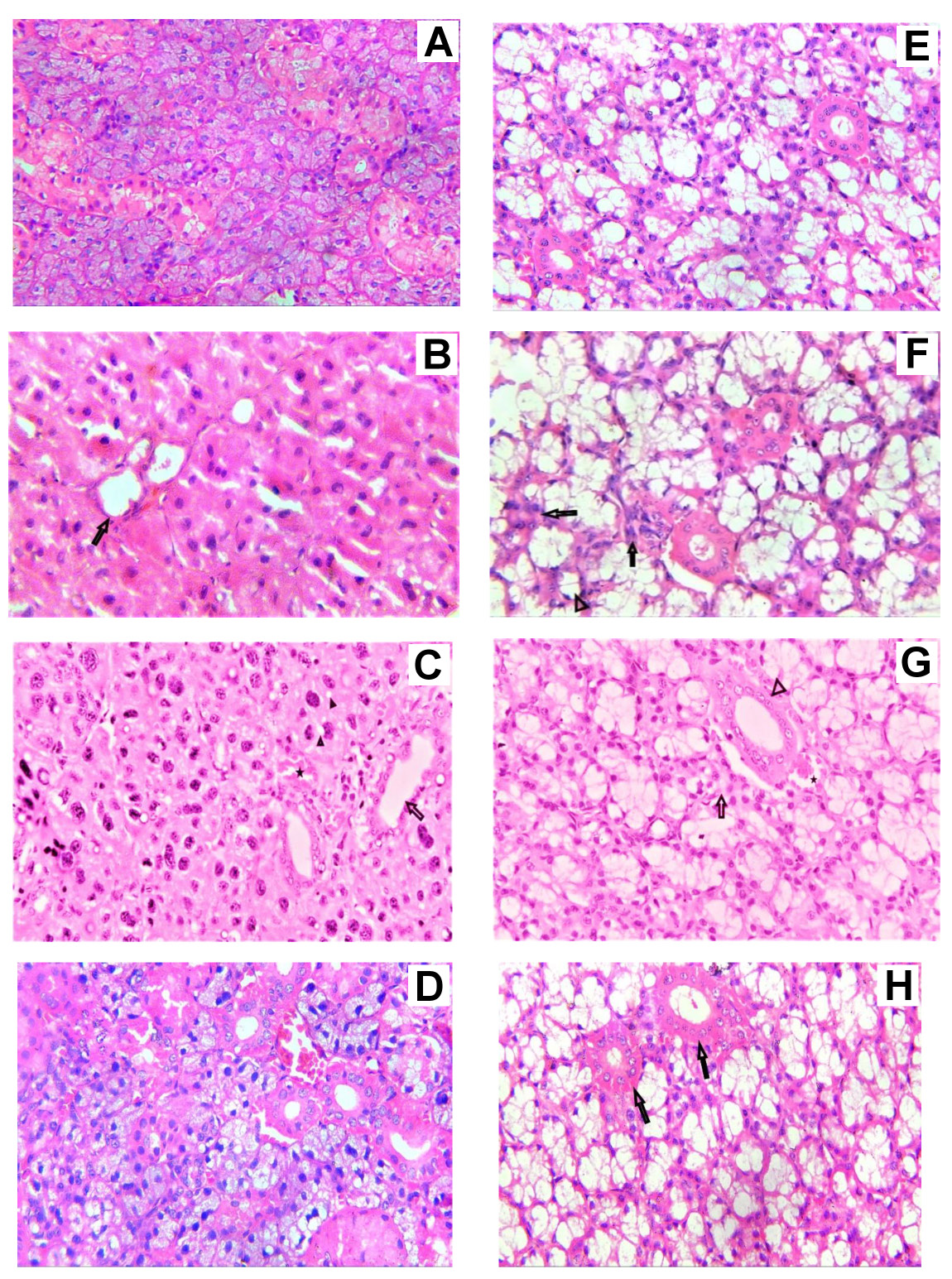
In group I, the parotid glands revealed a normal architecture of serous acini with pyramidal cells and a rounded basally located nucleus arranged around a small lumen (Figure 3A). The evaluation of the mandibular mucosal glands revealed the presence of typical mucous acini with oval, basally located nuclei encircling a broader lumen. The intralobular ducts of both glands were lined with columnar cells with centrally placed, large, rounded nuclei (Figure 3E).
In group II, the examination of parotid sections revealed loss of the normal acinar architecture with signs of deterioration. The nuclei were irregularly arranged, with variable sizes and shapes (Figure 3B,C). Mandibular mucosal gland sections demonstrated atrophy of the acini, and some showed loss of architecture. The intralobular ducts of both glands revealed degenerative changes and loss of normal structure (Figure 3F,G).
In group III, the sections of both glands revealed improvement in acinar structure. Additionally, the intralobular ducts re-established their normal architecture (Figure 3D,H).
Immunohistochemical evaluation
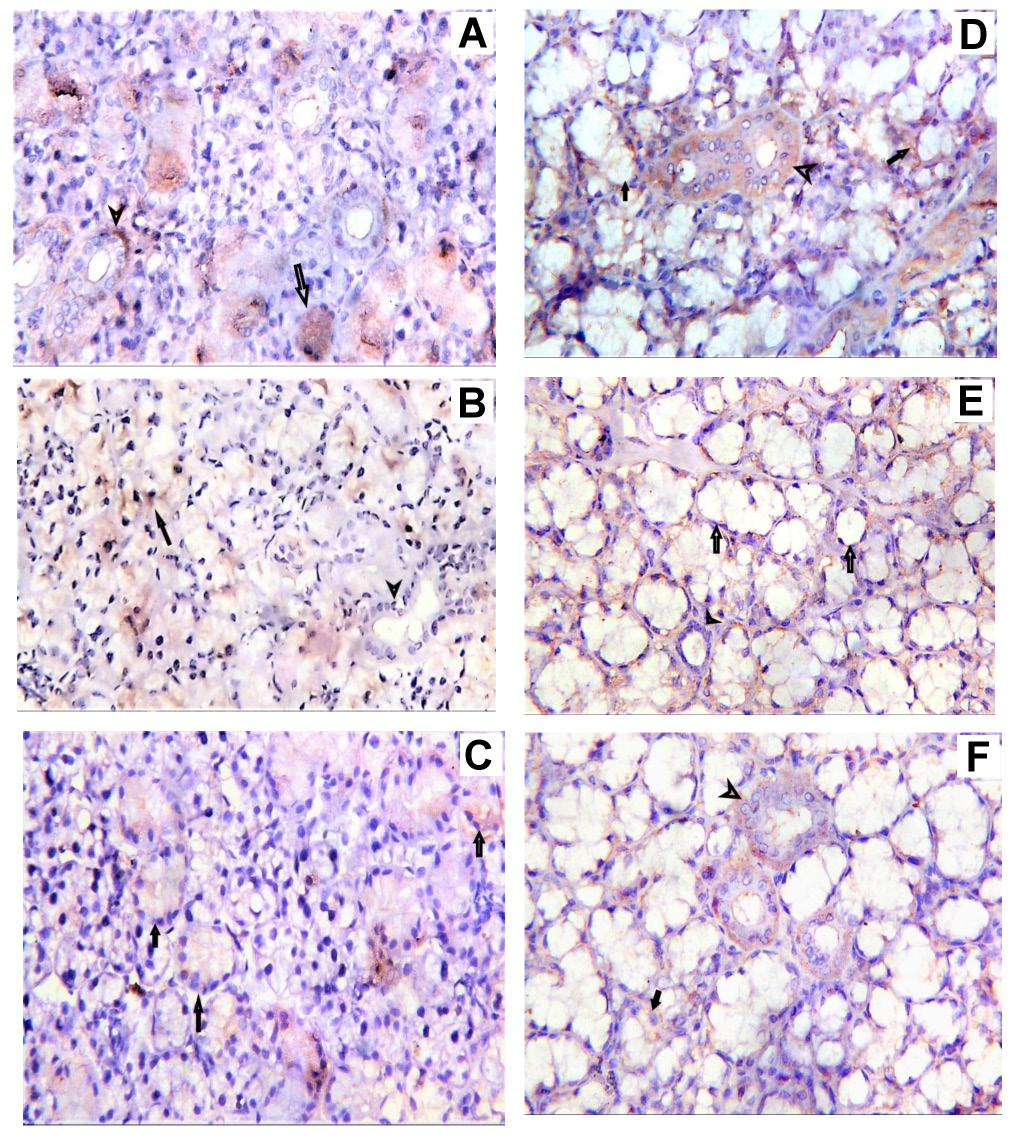
In group I, positive cytoplasmic staining was observed in the cells of both acini and ducts in the parotid and mandibular mucosal glands. In group II, a weak positive reaction was reported. In group III, acinar cells exhibited moderate positivity, which was lower than that reported in group I. Ductal cells presented stronger positive staining compared to group II (Figure 4). The expression of VEGF in the parotid and mandibular mucosal glands in all groups is presented in Table 2.
The results of one-way ANOVA and the post hoc LSD test for VEGF expression in the parotid and mandibular mucosal glands in all groups are presented in Table 3 and Table 4.
Comet assay analysis
The comet assay was conducted to examine the degree of cellular DNA damage in the parotid and mandibular mucosal gland cells in all groups. In group I, the majority of cells were observed as uniform discs, characterized by low tail moment, short tail length and a low percentage of tail DNA.
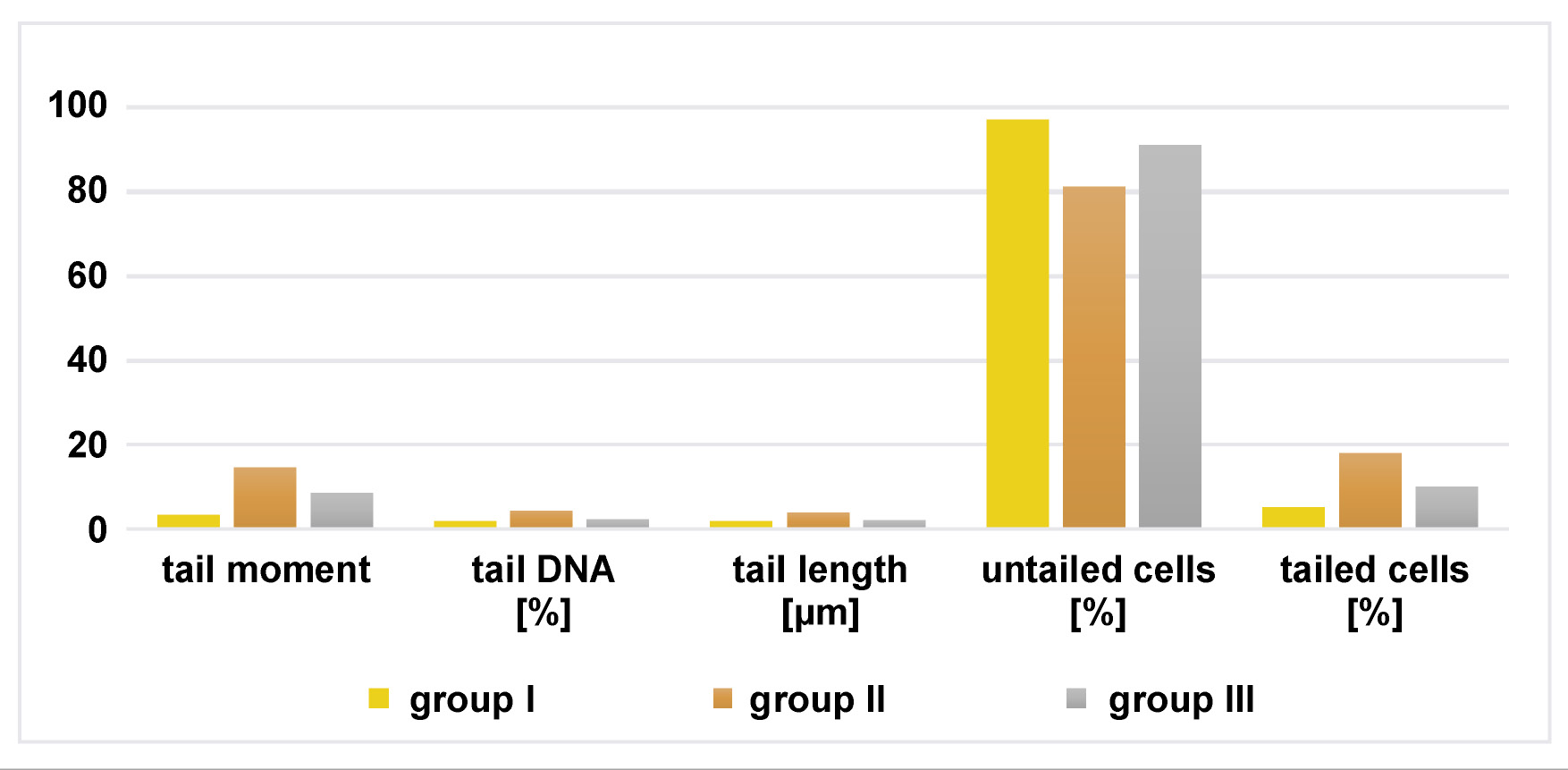
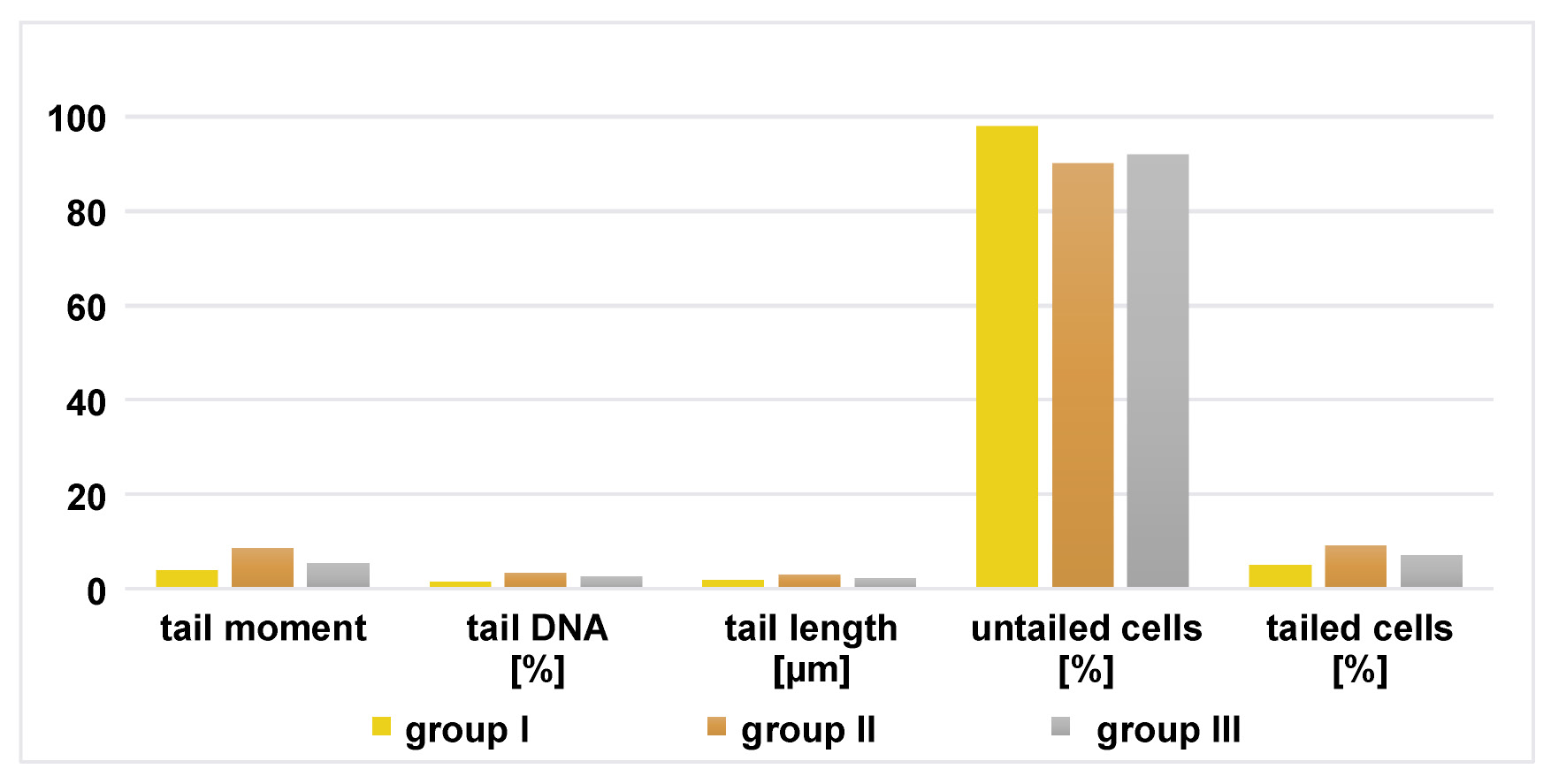
In group II, in both glands, the percentage of tailed cells, tail length, tail moment, and the percentage of tail DNA were significantly increased in comparison to group I. In group III, in both glands, the tail moment, tail length and the percentage of tailed cells were markedly reduced compared to group II (Figure 5, Figure 6).
The mean values of comet assay parameters were calculated for the parotid and mandibular mucosal glands in all groups, including the percentage of tail DNA, tail moment, tail length, and the percentage of tailed and untailed cells (Figure 7, Figure 8).
The results of comet assay in the parotid and mandibular mucosal glands for the 3 groups are presented in Table 5 and Table 6.
Discussion
The current study investigated the efficacy of chia seeds in restoring the architecture and histological integrity of the parotid and mandibular mucosal glands in rats with STZ-induced diabetes. Streptozotocin is widely used for the experimental induction of diabetes. Streptozotocin-induced diabetes in rats provides a reliable and well-established model for studying the deleterious effects of diabetes on multiple systems.36
The diabetic status induced by the administration of STZ was confirmed by elevated levels of serum glucose, nitric oxide and MDA. The administration of a single dose of STZ initiates an autoimmune process leading to the destruction of the Langerhans islet beta cells, resulting in the development of clinical diabetes within a short period (2–4 days) and the induction of oxidative stress.37
In the current study, the parotid gland specimens in group II demonstrated loss of normal acinar architecture with degenerative changes. The nuclei were irregularly arranged, with variable sizes and shapes. The intralobular ducts revealed deterioration and loss of normal structure. These findings are consistent with results of study by Cui et al., who reported serous acinar cell distortion and disintegration with reduced secretory granules, vacuolization and leukocyte invasion in the salivary glands of diabetic rats.38 In the present study, the observed changes in blood vessels may be attributed to the inflammatory processes associated with DM, as stated by Ror et al.39
In group II, the mandibular mucosal gland sections demonstrated acinar atrophy with loss of normal architecture, as well as deterioration and structural disruption of the intralobular ducts, which is in agreement with the findings of Stewart et al., who confirmed that diabetes is associated with hypofunction and atrophy of the parotid and mandibular mucosal salivary glands in diabetic rats due to reduced nitric oxide synthase activity, which is important for the proper functioning of salivary glands.40 These findings may be explained by the deleterious effects of chronic hyperglycemia, which promote oxidative stress.41 Oxidative stress results from multiple pathways, including glucose autoxidation, activation of the polyol pathway, and the formation of advanced glycation end-products (AGEs).42 Other circulating factors reported to be elevated during the course of diabetics, namely free fatty acids and leptin, are also associated with the increased production of ROS.43
Krippeit-Drews et al. explained the mechanisms of oxidative stress-induced cell injury through both indirect and direct pathways.44 Indirect pathways include the release of calcium from intracellular stores and its influx across the plasma membrane. The increased number of cytosolic calcium ions activates enzymes responsible for cellular damage, including phospholipases, proteases, endonucleases, and adenosine triphosphatases (ATPases), thereby accelerating ATP reduction. This ATP depletion causes structural disruption of protein synthesis. Moreover, ATP reduction impairs the activity of the plasma membrane energy-dependent sodium pump, leading to intracellular aggregation of Na+ and loss of K+, resulting in cell enlargement and dilation of the endoplasmic reticulum.44 Hyperglycemia may also impair the function of sodium–glucose cotransporters and aquaporins, affecting cellular water balance.45
Conversely, direct pathways include peroxidation of polyunsaturated membrane lipids, which are susceptible to attack by oxygen-derived free radicals. Lipid radical interactions generate peroxides that compromise the integrity of cellular and nuclear membranes. This permits oxygen radicals to attack chromatin, leading to DNA disintegration. Additionally, free radicals react with thymine in nuclear and mitochondrial DNA, causing single-strand breaks. This DNA damage contributes to cellular aging and apoptosis.46
Whole chia seeds were administered in the form of flour since the particle size directly influences digestion and metabolic processes.47 Furthermore, whole seed flour retains all structural components of the grain, including endosperm, bran, germ, and coat.48 Moreover, grinding enhances the bioaccessibility of bioactive antioxidants, such as phenolic acids, thereby increasing their potential health benefits.49
In group III, which was subjected to diabetic induction and treated with chia seeds, the parotid and mandibular mucosal glands showed improvement in acinar architecture and intralobular duct structure. These findings indicate that chia seeds might have an ameliorative effect on salivary glands affected by diabetes-induced degeneration. Moreover, Enes et al. reported similar findings regarding the effects of chia seeds on glucose metabolism in both in vitro and in vivo studies.34 The authors demonstrated that chia flour and chia oil decreased adiposity; however, only chia oil was able to improve glucose tolerance and restore the energy fuel system in the liver of rats fed a high-fat high-fructose (HFHF) diet. In addition, chia hydroalcoholic protein extract (CHPE) decreased the mRNA expression levels of gluconeogenic enzymes.34
In addition, de Paula Dias Moreira et al. stated that the inclusion of chia flour or chia oil in an unbalanced diet rich in saturated fat and fructose promoted beneficial effects on liver health by reducing metabolic disturbances through modulation of different metabolic pathways.50 Chia flour and chia oil restored antioxidant system activity and enhanced fatty acid oxidation. Furthermore, chia flour modulated lipogenesis, whereas chia oil improved blood glucose levels, triglyceride (TG) levels and body weight.50
Chia seeds contain a wide range of antioxidant compounds, including vitamins C and E, carotenoids, phenolic compounds, and polyphenols such as flavonoids.51 The flavonoid group with antioxidant activity includes flavones, flavonols, isoflavones, catechins, and chalcones. Micronutrients present in plants, such as vitamins A, C and E, folic acid, carotenoids, anthocyanins, and polyphenols, can scavenge free radicals and may serve as natural alternatives to synthetic antioxidants.52
Furthermore, chia is one of the most notable natural sources of omega-3 fatty acids, which inhibit the production of prostaglandins and pro-inflammatory cytokines, thereby modulating the inflammatory response and indirectly reducing cellular damage.53 Additionally, the high levels of α-linolenic acid support the function of the epithelial barrier by maintaining the integrity of intercellular junction proteins.54
In a study conducted by Enes et al., the extraction of chia seeds into chia oil was shown to significantly reduce blood glucose levels.55 The consumption of chia seeds has been associated with the restoration of lipids and membrane fatty acid translocase (FAT/CD36) in the plasma membrane, as well as with enhanced mitochondrial function and beta-oxidation.56 The distribution of lipids may improve glucose tolerance by reducing body adiposity, an effect that has been associated with α-lipoic acid. Alpha-lipolic acid acts as an activator of adenosine monophosphate-activated protein kinase (AMPK). The activation of AMPK reduces blood glucose levels by promoting the phosphorylation of Akt substrate of 160 kDa (AS160), a protein active in glucose transporters, thereby enhancing insulin-dependent glucose uptake.57
A study conducted by Hassan Mahmoud et al. further supported the antioxidant activity of chia seeds.26 The authors investigated the impact of low-level laser therapy (LLLT) and chia seeds on the alleviation of photoreceptor aberrations in experimental diabetic retinopathy (DR) and reported that the association between LLLT and the antioxidant action of chia seeds accelerated the recovery of retinal photoreceptors. They demonstrated that in DR, the production of ROS is elevated during the early stages of diabetes, leading to structural damage to mitochondria in photoreceptors. Additionally, persistent hyperglycemia results in the excessive production of ROS, continuous mitochondrial damage and elevated oxidative stress. Chia seeds reduced oxidative stress and hastened photoreceptor layer (PRL) mitigation, likely due to their antioxidant properties.26
Vascular endothelial growth factor has a pleiotropic role in tissue repair via angiogenesis, re-epithelialization and maintenance of extracellular matrix balance.58 In group II, the expression of VEGF in acinar and ductal cells was reduced compared to groups I and III, which may be attributed to the adverse effects of diabetes on vascular function, including increased endothelial permeability to macromolecules and impaired endothelial proliferation.59 Radwan et al. also investigated the antidiabetic effect of bee venom compared with bone marrow mesenchymal stem cells on the rat tongue using VEGF as an indicator of regeneration. Comparable results were obtained, as VEGF expression in the diabetic group was lower than in the treated groups.60 Diabetes also disrupts the balance between cell proliferation and apoptosis.61 Studies on cutaneous wound healing in diabetic models have shown delayed re-epithelialization, absence of growth factors and decreased angiogenesis.62
The alkaline comet assay is utilized as a conventional technique for the identification of DNA damage in genotoxicity testing and human biomonitoring.63 Furthermore, it is an accepted method in ecotoxicology and environmental monitoring for the study of various animal and plant species.64 Hyperglycemia in DM increases the production of ROS, thus enhancing endothelial cell apoptosis in vivo and in vitro, promoting free radical formation and reducing antioxidant capacity due to glycation and lipid oxidation reactions.65 Oxidative modifications to DNA occur due to alterations of nucleotide bases and sugars, as well as the formation of cross-links, leading initially to base oxidation and fragmentation, followed by single- and double-strand breaks, interstrand and intrastrand cross-links, and eventually DNA–protein cross-links and sugar fragmentation products.66
In the current study, group II of both glands presented increased DNA damage, as evidenced by greater DNA tail length, higher percentage of tail DNA, and increased tail moment (defined as the product of tail length and the fraction of total DNA in the tail) compared to group I. In group III, DNA damage was markedly reduced following treatment with chia seeds compared to group II, indicating that the extent of oxidative stress correlates with the size of comet tail. These findings confirm the ability of chia seeds to ameliorate the deleterious effects of DM on salivary glands, as reflected by the improvement in acinar and ductal architecture. Moreover, the phenolic compounds present in chia seeds include flavonols and phenolic acids, such as myricetin, quercetin, kaempferol, and caffeine acid.67 These compounds act as primary and synergistic antioxidants, contributing to the overall antioxidant activity of chia seeds. Quercetin, in particular, is a powerful antioxidant that inhibits the oxidation of fats, proteins and DNA, and exhibits stronger antioxidant activity compared to other flavonoids.68
Limitations
The present research was conducted using an experimental animal model. Although such model provides invaluable insights, findings of the study cannot be directly extrapolated to humans. Moreover, the study did not include the monitoring of blood glucose levels before, during and after the administration of chia seeds.
Conclusions
The immunohistochemical and genotoxic results obtained from the present study demonstrated that diabetes induction leads to significant structural changes in the parotid and mandibular mucosal glands in rats, primarily through oxidative DNA damage. Treatment with chia seeds ameliorated these changes, likely due to their antioxidant properties. Chia seeds showed promising effects in reducing oxidative stress. However, further studies are required, including investigations with different treatment durations and dosages, to support the use of chia as a dietary supplement in the management of DM. Further research should also evaluate long-term safety, identify bioactive compounds, and elucidate the underlying mechanisms. Additionally, studies should assess the effects of chia seeds on salivary gland function (flow rate and composition) to better understand their potential clinical relevance. The interactions between the various bioactive compounds of chia seeds, as well as the specific compound or synergistic effect responsible for their observed health benefits, remain unclear.
Ethics approval and consent to participate
The study was approved by the Institutional Animal Care at Faculty of Medicine, Mansoura University, Egypt (approval No. A 11070898). The research was conducted in accordance with the ARRIVE (Animal Research: Reporting of in Vivo Experiments) guidelines and regulations (https://arriveguidelines.org). The maintenance and care of the experimental animals adhered to the International Guiding Principles for Biomedical Research Involving Animals.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
Not applicable.
Use of AI and AI-assisted technologies
Not applicable.