Abstract
Background. Diabetes mellitus (DM) is a critical chronic metabolic disease. Several treatment modalities are currently under investigation. Both bee venom (BV) and bone marrow mesenchymal stem cells (BMSCs) can possibly offer an approach for treating type I diabetes.
Objectives. The aim of the present study was to investigate the mechanism underlying the anti-diabetic effect of BV as compared to BMSCs on the tongue mucosa of diabetic rats.
Material and methods. A total of 52 male albino rats were used in the current study. The rats were randomly assigned into 4 groups: group 1 (control); group 2 (streptozocin (STZ)); group 3 (BV-treated); and group 4 (BMSC-treated). Diabetes mellitus was induced via an intraperitoneal (IP) injection of STZ in the rats from groups 2, 3 and 4. Following the diagnosis of DM, the rats in group 3 were injected with a daily dose of 0.5 mg/kg of BV, while the rats in group 4 were treated with a single injection of BMSCs. All rats were euthanized after 4 weeks, and their tongues were dissected and divided into halves. The right halves of the tongues were utilized for the histological examination, followed by morphometric analysis. In contrast, the left halves were used to detect the local gene expression of transforming growth factor beta 1 (TGF-β1) and vascular endothelial growth factor (VEGF).
Results. Group 2 revealed marked disruption in the morphology of the fungiform and filiform papillae, and atrophic epithelial changes in both dorsal and ventral surface epithelium as compared to other groups. Group 4 showed a significantly larger number of taste buds, and a higher gene expression of TGF-β1 and VEGF as compared to groups 2 and 3. Additionally, BV and BMSCs effectively increased the thickness of dorsal and ventral surface epithelium as compared to group 2.
Conclusions. Treatment with BMSCs was associated with significant improvement in the morphology and number of lingual epithelial cells and taste buds in the tongues of diabetic rats as compared to BV-treated rats, which was due to the local upregulation of TGF-β1 and VEGF gene expression.
Keywords: bee venom, diabetes, streptozocin, bone marrow mesenchymal stem cells
Introduction
Diabetes mellitus (DM) is considered a critical chronic metabolic disease due to a prompt rise in the number of diagnosed patients, especially over the last 2 decades.1 Diabetes mellitus is characterized by high blood glucose levels, which, if not controlled efficiently, leads to end-organ damage within the genitourinary, cardiovascular and neurological systems.2 These characteristics make DM a condition that needs proficient and long-standing medical care in order to control elevated blood glucose levels and prevent complications.3 Several drugs used to treat DM have adverse effects, which necessitates a search for other modalities that could overcome these side effects, without imposing a financial strain on the patient.4
Bee venom (BV) is composed of different types of proteins, enzymes and non-peptide components. The proteins include melittin, apamin, adolapin, and mast cell degranulating (MCD) peptide. The enzymes found in BV include α-glucosidase, phosphatase, phospholipase B, phospholipase A2, and hyaluronidase. Additionally, non-peptide components, such as histamine, dopamine and norepinephrine, have been identified in BV.5 Due to its therapeutic effects, BV has been used as a drug in the treatment of many diseases, such as cardiovascular, neurological, hematological, musculoskeletal, and dermatological diseases.6, 7
The anti-diabetic effect of BV can be associated with melittin and phospholipase A2, a polypeptide and an enzyme that increase the secretion of insulin from pancreatic β-cells via the depolarization of the β-cell membrane.8 Taking into account the abovementioned properties, BV could be considered a therapeutic agent in the treatment of DM.
Bone marrow mesenchymal stem cells (BMSCs) are multipotent, self-renewing cell populations that exhibit a marked therapeutic potential because of their ability to differentiate into the 3 germ layer lineages. Furthermore, they can migrate toward the damaged sites when administered systemically. In damaged tissues, they can improve recovery, as they differentiate into cells specific to the tissue, and produce paracrine mediators and trophic factors that possess anti-apoptotic properties and stimulate cell proliferation.9, 10, 11
The BMSC therapy offers a cell-based approach for treating type I diabetes. These cells can differentiate into insulin-producing cells, and also show immunosuppressive activity, exerting ameliorative effects on injured tissues.12, 13
Diabetes mellitus is often related to prolonged or insufficient healing; this is attributed to the abnormal inflammatory response and a reduction in the production of growth factors, leading to impaired neovascularization and compressed collagen matrices.14
Transforming growth factor beta 1 (TGF-β1) is known to regulate the chemotaxis of immune and inflammatory cells, cellular differentiation, and induce the accumulation of extracellular matrix proteins.15 Indeed, defective TGF-β1 signaling contributes to delayed wound healing in diabetes.16 Vascular endothelial growth factor (VEGF) promotes tissue repair via increasing vascular permeability, as well as the proliferation and migration of the pre-existing endothelial cells.17
The present study aimed to evaluate and compare the potential anti-diabetic effects of BV and BMSCs against the histological and molecular changes in the tongue in streptozotocin (STZ)-induced diabetic albino rats.
Material and methods
Bee venom
The BV samples were collected from the colonies of Italian and Carniolan hybrid honeybees (Apis mellifera), using a BV collector (an electric shock device, VC-Starter kit; IGK electronics Ltd., Varna, Bulgaria) at the National Research Center, Cairo, Egypt. The dried BV material was transferred to a proper container and was solubilized in distilled water to reach a concentration of about 0.1 mg/mL.18
Bone marrow mesenchymal stem cells (BMSCs)
Bone marrow mesenchymal stem cells were isolated from the femora and tibiae of 6 Wistar donor rats (6-week-old, male, weighing 100 ±20 g). The isolation and propagation of BMSCs was conducted under aseptic conditions, 14 days before the experimental procedures, as previously described.19
The femora and tibiae were flushed with Dulbecco’s Modified Eagle Medium (DMEM) (Gibco-BRL, Gaithersburg, USA) supplemented with 10% fetal bovine serum (FBS) (Gibco-BRL), using an 18-gauge needle. The cells were cultured in a culture medium supplemented with 1% penicillin–streptomycin (Gibco-BRL) in an incubator, at 37°C in a humidified atmosphere (5% CO2). Upon reaching 80–90% confluence, the cultures were washed twice with phosphate-buffered saline (PBS) and the cells were trypsinized with 0.25% trypsin in 1 mM ethylenediaminetetraacetic acid (EDTA) (Gibco-BRL) at 37°C for 5 min. Then, the cells were centrifuged at 2,400 rpm for 20 min, re-suspended with a serum-supplemented medium and incubated in a Falcon® culture flask with a surface area of 50 cm2. On day 14, the adherent colonies of cells were trypsinized and counted.19
Culture confluence was monitored using an inverted light microscope (Olympus, Center Valley, PA, USA) with a digital camera (Nikon, Tokyo, Japan).
Characterization of BMSCs
Flow cytometry was performed to identify BMSCs. After blocking in 0.5% bovine serum albumin (BSA) and 2% FBS in PBS, 100,000 cells were incubated in the dark at 4°C for 20 min with the following monoclonal antibodies: FITC CD 90 (PN IM1839U; Beckman Coulter, Brea, USA); and PE CD 34 (PN IM1871U; Beckman Coulter). Mouse-isotype PE antibodies (Beckman Coulter) were used as controls (the dilution of all antibodies at 1:1,500). The cells were washed and suspended in 500 µL of the fluorescence-activated cell sorting (FACS) buffer, and analyzed using the Cytomics FC 500 flow cytometer with the CPX software, v. 2.2 (Beckman Coulter).
Animals
This study was approved by the Institutional Animal Care and Use Committee (IACUC) at Cairo University, Egypt (approval No. CU III F 74 18). This research was conducted in compliance with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines and regulations (https://arriveguidelines.org).
A sample size of 52 (13 per group) was estimated to be sufficient, assuming an effect size of 0.6, a power of 0.8, a two-sided hypothesis test, and a significance level of 0.05 for categorical and numerical data. Fifty-two healthy adult male albino rats, weighing about 150–200 g, with normal glucose levels, were used. All animals were housed in a sterile, controlled environment (room temperature of 25 ±5°C under a natural light, 12-hour dark/light cycle). They were fed with standard, ordinary, commercial rat pellets and were provided water ad libitum. The maintenance and care of the experimental animals conformed with the International Guiding Principles for Biomedical Research Involving Animals. The animals were randomly divided into 4 groups of 13 animals, using the Random Sequence Generator program (https://www.random.org).
Induction of experimental diabetes
Streptozotocin (Sigma-Aldrich, St. Louis, USA)20 was used for inducing type I diabetes by a single intraperitoneal (IP) injection of STZ (60 mg/kg b.w.) into fasted rats. The rats were considered diabetic when their random blood glucose reading was >300 mg/dL at 72 h of the STZ injection.21 Diabetes was allowed to stabilize in the STZ-injected rats for 1 week. The assessment of blood glucose levels was performed weekly in all diabetic rats until the end of the experiment. In all groups, the blood samples were obtained from the tail vein and the blood glucose level was expressed in mg/dL.
Animal grouping
The animals were randomly divided into 4 groups, 13 animals in each, occupying separate cages (Table 1). Group 1 (control) comprised normal healthy rats, which were given a physiological saline solution (0.9% NaCl). The veterinarian administered the STZ induction. Then, the STZ-treated rats were randomly divided into 3 groups (2–4). The rats from group 2 (the STZ group) received a single IP injection of STZ (60 mg/kg in freshly prepared 0.1 mol/L citrate buffer (pH 4.5)) and were left untreated. The diabetic rats from group 3 (the BV-treated diabetic group) were injected IP with a daily dose of 0.5 mg/kg of BV for 4 weeks.22 The diabetic rats from group 4 (BMSC-treated diabetic group) were administered a single intravenous (IV) injection of the previously cultured BMSCs at a dose of 1 million cells/mL in PBS.23
Animal sacrifice and tissue preparation
All rats were euthanized by an intracardiac overdose of sodium thiopental (80 mg/kg) after 4 weeks. The tongues were dissected into 3 parts: two halves of the anterior two-thirds; and the posterior one-third of the tongue.
The specimens from the right halves of the anterior two-thirds and the posterior one-third of the tongue were prepared for the histopathological examination. Serial sections of 5 μm of the tongue tissues were cut and subjected to the following:
– the fluorescence detection of the PKH26-labeled BMSCs in the unstained paraffin sections with the use of a fluorescence microscope; and
– the hematoxylin and eosin (H&E) staining for histological evaluation.
The specimens from the left halves of the anterior two-thirds were used to measure the expression of both TGF-β1 and VEGF.
The assessment steps were carried out blindly by the investigators.
Morphometric analysis
The specimens were examined using light microscopy (Leica, St. Gallen, Switzerland) under ×400 magnification. The data was obtained using the Leica Qwin 500 image analyzer computer system (Leica Biosystems, Cambridge, UK). Image analysis was done using the ImageJ software, v. 1.53d (https://imagej.net/ij).24 The image analysis system was used to assess the dorsal and ventral epithelial thickness, the thickness of each cell layer,25 and the area of the connective tissue papillae.26 For each criterion, 5 non-overlapping microscopic fields were randomly selected and evaluated. As previously described, the number of taste buds per circumvallate papilla was assessed.27
Quantitative real-time polymerase chain reaction
To analyze the mRNA levels of TGF-β1 and VEGF, the total RNA-containing genes were determined by the quantitative real-time polymerase chain reaction (qRT-PCR). RNA was isolated with the QIAzol lysis reagent (Qiagen, Venlo, the Netherlands). Complementary DNA (cDNA) was produced using a cDNA synthesis kit (Applied Biosystems, Waltham, USA), and qRT-PCR was performed using the StepOnePlus™ RT-PCR system (Applied Biosystems), following the standardized protocols. The expression of genes was normalized relative to the mean critical threshold (CT) values with the ΔΔCT method, using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA as an internal control. The primers for TGF-β1, VEGF and GADPH are listed in Table 2.
Statistical analysis
The values are presented as mean and standard deviation (M ±SD). The Kolmogorov–Smirnov test indicated that the data was normally distributed. The one-way analysis of variance (ANOVA) was used to assess differences between the groups, and Tukey’s post hoc test was applied when ANOVA yielded a significant difference. The significance level was set at p < 0.05. Statistical analysis was performed using the Minitab® software, v. 18.1 (https://www.minitab.com/en-us/support/downloads).
Results
Characteristics of BMSCs in the culture
The cultured BMSCs revealed a fibroblast-like morphology and they adhered to the tissue culture substrate within 24–48 h. They reached confluence within 7–14 days (Figure 1).
Fluorescence detection
The examination of the unstained paraffin sections with the use of a fluorescence microscope was performed to detect and track the PKH26-labeled BMSCs. The tongue specimens from group 4 injected with the PKH26-labeled BMSCs showed red fluorescent cells within the tongue tissue (Figure 2).
Characterization of BMSCs
The cells were characterized by flow cytometry through the use of surface markers. The results showed that most of the BMSCs were positive for CD90 and negative for CD34 (Figure 3).
Biochemical analysis
At 3 weeks, BMSCs significantly reduced blood glucose levels as compared to the STZ group. At 4 weeks, both BV and BMSC treatment were associated with a significant reduction in blood glucose levels as compared to the STZ group (Table 3).
Histopathological examination
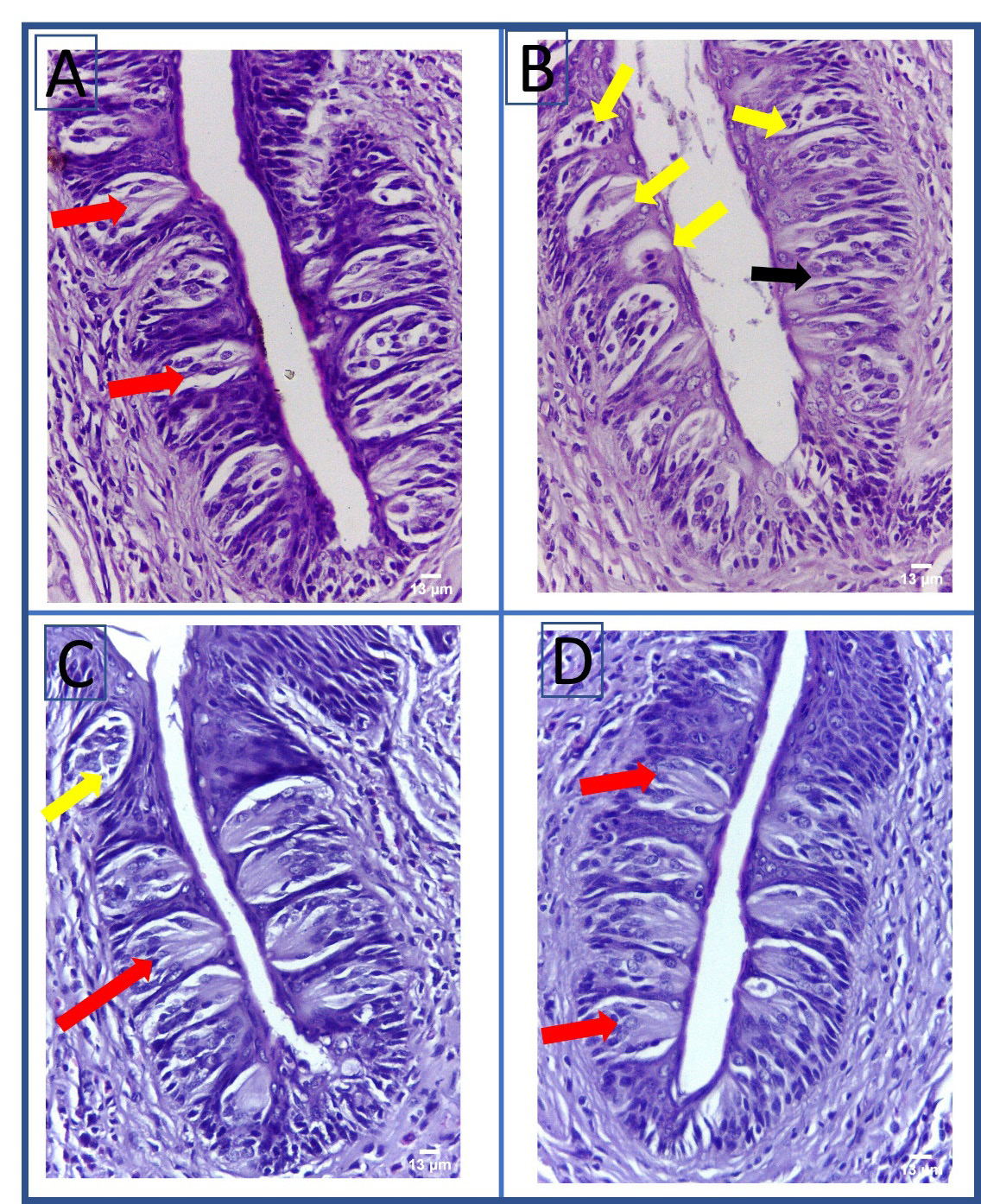
Upon the examination of group 1 (the control group), the fungiform papillae displayed a normal mushroom-like appearance with barrel-shaped taste buds and well-defined connective tissue (Figure 4). Normal thread-like filiform papillae with orthokeratinized epithelium and normal underlying connective tissue were detectable (Figure 5). The circumvallate papillae showed normal barrel-shaped taste buds (Figure 6). Dorsal and ventral surface epithelium revealed a normal thickness, was covered with normal orthokeratin and showed normal underlying connective tissue (Figure 7 and Figure 8).
Group 2 (the STZ group) showed disruption in the normal mushroom-like appearance of the fungiform papillae, and areas of a detached keratin layer and connective tissue degeneration. The fungiform papilla-associated taste buds revealed intracellular vacuolation and evidence of degeneration (Figure 4). The filiform papillae displayed atrophied, rounded tips, and a thin layer of keratin with areas of detached keratin. Some epithelial cells revealed hyperchromatic nuclei. The underlying connective tissue displayed evidence of degeneration (Figure 5). The circumvallate papillae showed multiple degenerated taste buds, while some taste buds presented the loss of their normal barrel-shaped outline (Figure 6). Dorsal surface epithelium revealed a reduced thickness and areas of detached keratin. In a few epithelial cells, hyperchromatic nuclei were detectable. There were areas of connective tissue degeneration and dilated blood vessels (Figure 7). The epithelium covering the ventral surface of the tongue showed epithelial atrophy, while the connective tissue showed areas of degeneration and dilated blood vessels (Figure 8).
Group 3, treated with BV, showed normal fungiform papillae with a mushroom-like appearance and normal barrel-shaped taste buds, with areas of connective tissue vacuolation (Figure 4). The examination of the filiform papillae revealed a thread-like shape, and multiple atrophied, rounded tips with areas of detached keratin (Figure 5). The circumvallate papillae showed few degenerated taste buds (Figure 6). Dorsal and ventral surface epithelium showed increased epithelial and keratin thickness as compared to Group 2, and areas of connective tissue degeneration (Figure 7 and Figure 8).
The examination of group 4, treated with BMSCs, showed mushroom-like fungiform papillae, with normal barrel-shaped taste buds and well-defined connective tissue (Figure 4). Thread-like shaped filiform papillae with few areas of detached keratin were detectable (Figure 5). Normal circumvallate papillae with normal barrel-shaped taste buds were evident (Figure 6). Dorsal and ventral surface epithelium showed an increase in the epithelium and keratin thickness as compared to groups 2 and 3, and areas of connective tissue degeneration (Figure 7 and Figure 8).
Morphometric analysis
The highest number of taste buds within the circumvallate papillae was detected in group 1, while the lowest was recorded in group 2, with a statistically significant difference between the groups (p < 0.05). Pairwise comparisons revealed a significantly higher number of taste buds in group 4 as compared to groups 2 and 3 (p < 0.05). Also, a significantly higher number was recorded in group 3 as compared to group 2 (p = 0.002) (Table 4).
Group 2 was associated with atrophic epithelial changes, with a significantly reduced thickness of both dorsal and ventral surface epithelium as compared to other groups (p < 0.05). Groups 3 and 4 effectively increased the epithelial thickness, which showed significantly higher values in both dorsal and ventral surface epithelium as compared to group 2 (p < 0.05). Regarding the dorsal and ventral epithelial thickness, a higher mean value was reported for group 4 as compared to group 3. However, the difference was not statistically significant (Table 4).
A reduction in the dorsal epithelial thickness in group 2 as compared to other groups was associated with a significant reduction in the thickness of the basal cell, polyhedral cell, granular cell, and keratin layers (p < 0.05) (Table 4).
Both group 3 and group 4 showed a significant increase in the thickness of polyhedral cell, granular cell and keratin layers as compared to group 2 (p < 0.05), whereas the thickness of the basal cell layer showed a significant increase only in group 4 as compared to group 2 (p < 0.05). A higher mean value was recorded for the epithelial layer thickness in group 4 as compared to group 3; however, the difference was not statistically significant except for the polyhedral cell layer thickness (p < 0.05) (Table 4).
Group 2 showed a significant reduction in the area of the connective tissue papillae (p < 0.05). Group 3 had significantly increased connective tissue papillae as compared to groups 2 and 4 (p < 0.05). An insignificantly higher mean area of the connective tissue papillae was detected in group 4 as compared to group 2 (p < 0.05) (Table 4).
TGF-β1 and VEGF gene expression
Group 2 showed a significant reduction in TGF-β1 and VEGF expression as compared to other groups (p < 0.05). Group 4 showed a significant increase in TGF-β1 and VEGF expression as compared to groups 2 and 3 (p < 0.05) (Table 5).
Discussion
Diabetes mellitus is a metabolic disorder resulting in multiple complications, including soft tissue abnormalities in the oral cavity. In the present study, we investigated the possible therapeutic effects of BV and BMSCs on the degenerative changes affecting the tongue due to the induction of DM in rats.
Streptozotocin was chosen for DM induction in rats, as it provides a permanent and stable diabetic model. It can induce type I diabetes through the rapid destruction of β-cells in the pancreas.28 In the current study, STZ-induced DM in rats was associated with atrophic changes to the lingual mucosa, with a reduction in the thickness of both dorsal and ventral surface epithelium, the atrophy of the lingual papillae, and a reduction in the number of taste buds per circumvallate papilla. The atrophy of the underlying connective tissue with a significant reduction in the connective tissue papillae was also noticed.
The effect of uncontrolled DM on rats’ tongue epithelium and papillae has been previously reported. Streptozotocin-induced DM in rats has been associated with alterations in the distribution and morphology of the fungiform and filiform papillae, with areas of epithelial desquamation,29, 30, 31 in addition to a significant reduction in the height and width of the filiform papillae.32 Diabetes mellitus has also been associated with the atrophy and a decreased thickness of dorsal surface epithelium,32, 33, 34 and the atrophy of ventral surface epithelium.34 Lingual epithelial atrophy was observed in rats born to diabetic mothers.35 Epithelial atrophy was also noticeable in the buccal mucosa of diabetic rats.36
In the present study, the circumvallate papillae were impacted by DM. A significant reduction in the number of taste buds per circumvallate papilla has been reported in previous studies.27, 37 These changes were attributed to a DM-associated reduction in the innervation of taste buds.27 Additionally, rats with induced DM were prone to increased apoptosis of circumvallate papilla taste buds, which was associated with the downregulation of Bcl-2, the upregulation of Bax, and increased activation of caspase-9 and caspase-3.37
Diabetes mellitus has also been linked to the atrophy of the lamina propria underlying the lingual mucosa. A decrease in the height and cross-sectional area of the connective tissue papillae of the dorsal surface of the tongue in rats with induced DM has been previously reported.26, 31 In addition to the atrophied gingival lamina propria,25, 38 DM was also linked to an increased incidence of Candida albicans and the thickening of bacterial colonies on the dorsal surface of the tongue.29
On the other hand, some reports demonstrated that DM was associated with epithelial hypertrophy.25, 38 The thickness of the gingival epithelium, and of the prickle cell, granular cell and keratin layers was significantly higher in rats with induced DM as compared to the control group.25 A statistically significant increase in the thickness of the gingival epithelium, and of the basal, prickle cell, granular, and keratin layers was also observed.38 This indicates that regional variations in the oral tissues can affect the mucosal response to chemically induced DM in rats.
The atrophic effect of DM on the lingual mucosa can be traced back to the diabetes effect on epithelial cells, as DM has been associated with reduced keratinocyte proliferation39 and increased cellular apoptosis.40 Reduced cell proliferation, in addition to increased cellular apoptosis, can result in epithelial atrophy. The effect of DM on cellular functions has been attributed to high blood glucose levels, the accumulation of advanced glycation end-products (AGEs), increased tissue hypoxia, and increased levels of reactive oxygen species (ROS) in DM.41, 42, 43 Diabetes mellitus can also cause systemic inflammation and a chronic inflammatory infiltrate in the oral connective tissues, which can negatively affect the integrity and function of the oral mucosa.44, 45, 46, 47 Additionally, diabetic microangiopathy may lead to subsequent connective tissue degeneration and mucosal atrophy.25, 26, 48 It has been observed that DM is also linked to an increased expression of p53 in rats33 and an increased expression of p16.32
The positive effect of BV on the lingual mucosa observed in the current study reflects its potent biological properties. Bee venom shows anti-inflammatory,49 antibacterial50 and antioxidant effects.51 In addition, BV has demonstrated anti-cancerous effects52, 53 and anti-obesity effects.53 It also has an anti-diabetic effect, as it can lower a blood glucose level, increase the secretion of insulin53 and enhance diabetic wound healing.51, 53, 54, 55, 56 The anti-inflammatory activity of BV could be attributed to melittin, its main constituent, as melittin inhibits the enzymatic activity of phospholipase A2.51 The phospholipase A2 enzyme is released in severe inflammatory disorders and causes tissue damage.
Bone marrow mesenchymal stem cells effectively improved the morphology of the lingual papillae, increased the number of taste buds per circumvallate papilla and effectively reversed DM-associated epithelial atrophy. Similar to our findings, BMSCs effectively reversed atrophic changes in the fungiform and filiform papillae,30, 34, 57 lingual mucosa,34 and increased the ventral epithelial thickness in rats with STZ-induced DM.34 Bone marrow mesenchymal stem cells were also reported to improve the morphology of both the circumvallate and foliate papillae, and restore their taste buds in rats with STZ-induced DM.58 The regenerative effect of BMSC on the lingual mucosa can be attributed to the ability of the cells to release different signaling molecules, including growth factors, cytokines and chemokines, in addition to their anti-inflammatory effect.59
The ability of BMSCs to restore the lingual mucosa in diabetic rats can be partly attributed to their angiogenic effect. Bone marrow mesenchymal stem cells can induce angiogenesis in diabetic wounds through the release of pro-angiogenic factors, including hypoxia-inducible factor (HIF), VEGF, angiopoietin, and erythropoietin,60, 61 in addition to their ability to differentiate into endothelial cells.61 They can also secrete multiple growth factors, including epidermal growth factor (EGF), insulin-like growth factor 1 (IGF-1) and platelet-derived growth factor (PDGF), important for the chemotaxis and function of cells responsible for diabetic wound healing.62, 63 The cells also have immunomodulatory properties. They can interact with cells of the innate and adaptive immune systems to downregulate pro-inflammatory cytokines, including interleukin 1 beta (IL-1β), tumor necrosis factor alpha (TNF-α) and IL-6, and to upregulate anti-inflammatory cytokines – IL-10 and prostaglandin E2 (PGE2).64 Bone marrow mesenchymal stem cells can also increase the migration, proliferation and function of keratinocytes in diabetic wounds, and induce re-epithelialization.63, 65 Taken together, the ability of BMSCs to stimulate angiogenesis and keratinization, and their immunomodulatory properties can explain their potential to reverse the diabetes-associated atrophic changes observed in the lingual mucosa of rats in the current study.
In this study, the levels of TGF-β1 and VEGF were investigated to define their role in enhancing the regeneration of the tongue following BV and BMSC treatment in diabetic rats. Treatment with BV and BMSCs resulted in the upregulation of both TGF-β1 and VEGF in comparison with the levels observed in the diabetic rats in group 2.
The molecular mechanisms underlying the regeneration of damaged tongue tissues following BV treatment in group 3 may be related to the ability of BV to upregulate TGF-β1, which stimulates the migration of keratinocytes and increases integrin expression.66 Transforming growth factor beta 1 can induce collagen expression.67 Also, BV enhances the expression of VEGF, which in turn stimulates local angiogenesis through mobilizing and recruiting bone marrow-derived endothelial progenitor cells, thereby decreasing impaired healing in diabetic rats.51
According to Kwon et al., the regenerative potential of BMSCs in tongue tissues in diabetic rats appeared to provide better results when compared to the untreated animals, which was explained by the local upregulation of cytokines and growth factors.62 Bone marrow mesenchymal stem cells increase the expression of TGF-β1 moderately and of VEGF markedly, which in turn enhances the inflammatory response, and induces the recruitment and proliferation of cells, required for repairing damaged tissues. Additionally, the immunoblotting analysis revealed an increased expression of neovascularization-related genes, such as TGF-β1 and VEGF, in a BMSC-treated mouse burn injury model.68 In the present study, group 2 (diabetic rats) displayed the lowest VEGF levels, which might be due to the inability of diabetic rats to properly upregulate VEGF expression in response to hypoxia.20
Conclusions
Diabetes mellitus exerted detrimental effects on rat tongues. Therapy with BV and BMSCs ameliorated the damaging effects of DM by upregulating the expression of TGF-β1 and VEGF. However, BMSC therapy promoted better results in regenerating the diseased tissues.
Ethics approval and consent to participate
This study was approved by the Institutional Animal Care and Use Committee (IACUC) at Cairo University, Egypt (approval No. CU III F 74 18). This research was conducted in compliance with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines and regulations (https://arriveguidelines.org). The maintenance and care of the experimental animals conformed with the International Guiding Principles for Biomedical Research Involving Animals.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
Not applicable.